Sex and gender
Our main focus here will be development of the reproductive system in the female and male, together with some of the variations that can occur. The metaphor to have in mind is that of a single path that everyone follows to begin with, and then from time to time the path splits into two alternative paths. At each fork we have to make a choice about which way to go, and having made that choice we have to accept that there can be no turning back at a later time to try the other path.
At first sight, it seems fairly straightforward to categorise people as either female or male. Having made that distinction, there will then be expectations about physical appearance, behaviour, and roles within society. Different societies might set different rules for each sex. However, the development of the biological make-up of a person is a complex multi-dimensional process composed of many steps, and added to that will be the many personal and cultural influences that together will shape the complete gender of the person. Traditionally, societies have taken a polarised view of sexuality and gender in that an individual is considered to be either one sex or the other, and legal frameworks generally reflect this polarisation. Increasingly that compartmentalism is being challenged and there is a realisation that the sex and gender of an individual occupies a position somewhere along a spectrum between the two poles of femininity and masculinity, and may also exist on a side arm branching away from the linear spectrum. Gender has become more fluid, and people who are categorised as either male or female at birth may feel at odds with the assigned gender and later transition to the gender that they feel they are.
There are several key branch points along the pathways to becoming reproductively-able males or females. The first is the establishment of genetic sex, followed by differentiation of the gonads (the general name for organs that produce gametes) into either ovaries or testes, then the selection of the correct internal ducts and associated reproductive structures accompanied by appropriate differentiation of the external genitalia. As birth approaches, the developing brain is influenced by the balance of hormones circulating in the baby's blood. Further development occurs after birth and through puberty to produce the adult form.
genetic sex
The development of biological sex begins at conception when the fertilising sperm arrives with either an X chromosome or a Y chromosome in the haploid genome that it contributes to the ovum. If the sperm carries an X chromosome, the new individual will be female (44 autosomes + XX sex chromosomes), and if it carries a Y chromosome, the individual will be genetically male (44 + XY).
After conception, there follows a period of 6 weeks during which female and male embryos develop along identical pathways, and cannot be distinguished on morphological grounds alone. Only a chromosomal analysis at this time could distinguish the sex. This is called the indifferent period.
gonadal sex
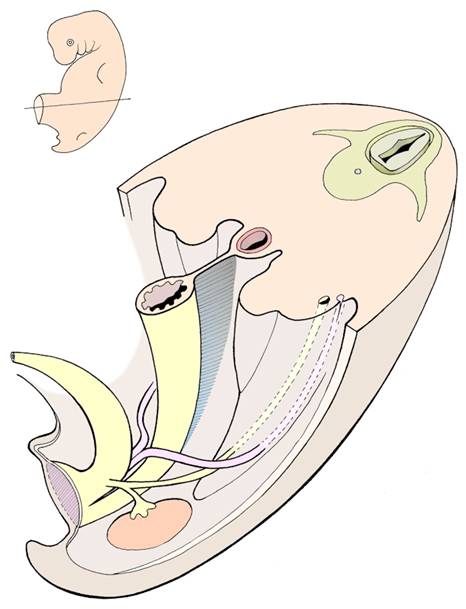
The rudimentary gonads arise as swellings on the posterior wall of the embryonic abdomen alongside the mesonephric ridges. Their initial development is the same in both sexes. Then an event of great significance occurs - some special cells called primordial germ cells migrate into the rudimentary gonads, having started their journey from the wall of the yolk sac. These cells will later give rise to the gametes after passing through two meiotic cell divisions and undergoing structural changes. (It is intriguing to speculate on why and how these important cells are set aside for their future role at such an early stage in development.) In genetic females, ovarian development takes place, with most of the changes occurring in the outer (cortical) regions of the gonad. In the male, it is the inner medulla of the gonad which shows greater development with formation of the seminiferous tubules and rete testis. The genetic information carried by the Y-chromosome plays a key role in stimulating testicular differentiation.
genital ducts
During the indifferent period, two duct systems form inside the embryonic abdomen. Later, only one duct system will be retained, and the other regresses. Which system is retained will depend on the genetic sex of the embryo and whether the gonads are developing as ovaries or testes. The two parallel systems are:
When the urorectal septum develops, it subdivides the cloacal membrane into an anal portion and a urogenital portion. Thus, the original coacal folds become modified to form the anal and urogenital folds respectively. (The intervening region, where the urorectal septum fuses with the cloacal membrane, will later differentiate into the clinically important perineal body.)
Soon after this stage is reached in the 6th week, the urogenital and anal membranes begin to perforate. From now on the development of the external genitalia will diverge along different pathways, depending on the genetic sex of the embryo, differentiation of the gonads into either ovaries or testes, and changes in circulating hormones. The main difference that can be seen between the two sexes in the following weeks as the external genitalia form is that the swellings in the urogenital region of the male embryo become fused with each other while in the female they remain separated.
In the female embryo, the genital swellings enlarge to form the labia majora, and the urethral folds become the labia minora. Both pairs of folds remain unfused. The genital tubercle differentiates - with the help of nearby tissues - into the clitoris. The short urethra opens directly into the vulval cleft, and the vagina - after canalisation - also communicates with this cleft, although its entrance is partially guarded by the membranous hymen.
In the male embryo, the genital swellings enlarge and fuse caudally to form the scrotum. Later, the testes will descend from the abdomen into the scrotum. The urethral folds approach each other in the midline and fuse, forming the shaft of the penis, and enclosing the penile part of the urethra. The genital tubercle forms the glans penis, which at first is a solid structure. A lumen then develops from the tip of the glans and extends back until it joins the penile urethra. Thus, the urethra in the male has a composite origin, receiving a contribution from the urogenital sinus, being added to by closure of the urethral folds, and being completed by canalisation of the glans penis.
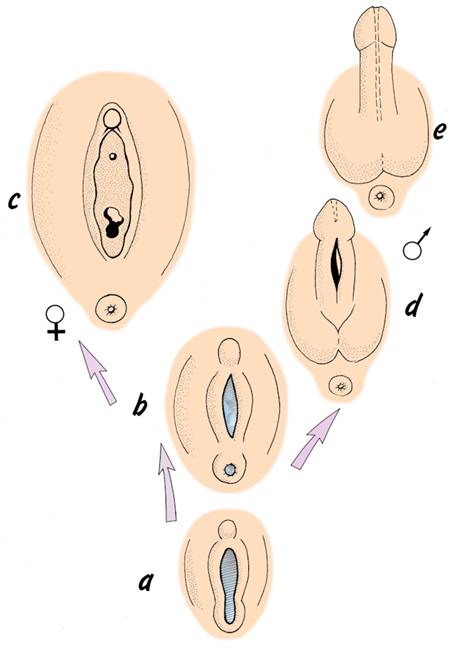
External genitalia - (a) during indifferent period,
(b) contact of urorectal septum with cloacal membrane,
(c) labia minora and majora of female perineum,
(d) formation of scrotum and shaft of penis,
(e) canalisation of glans penis to complete urethra.
descent of the gonads
The gonads develop on the posterior wall of the abdomen, but then in embryos of both sexes 'descend' to a more caudal position. The male gonads descend further than their female counterparts - the testes leave the abdominal cavity and enter the scrotum, where the slightly lower temperature will be more conducive to sperm-formation after puberty. The ovaries descend into the pelvic region and finish their migration alongside the uterus.
The descent of the gonads is controlled by a structure called gubernaculum and aided by elongation of the abdominal region of the embryo and later fetus. The gubernaculum is a fibromuscular strand extending from the caudal part of the gonad down into the genital swelling of the same side. It has been proposed that this structure is actively contractile, but not everyone agrees with this view. In the female embryo, the gubernaculum lies alongside the developing uterus in its mid-region and becomes attached to it. This subdivides the gubernaculum into two portions: one extending from the ovary to the uterus (which later becomes the ligament of the ovary), and the other from the uterus to the genital swelling (later the round ligament of the uterus). The result is that the ovary normally descends only as far as the pelvic region and comes to rest alongside the uterus on the posterior aspect of the broad ligament. Very rarely an ovary might descend into a labium majus.
In the male embryo, the gubernaculum does not become subdivided. The testis moves down the posterior abdominal wall and then through the anterior abdominal wall into the scrotum. As it passes through the abdominal wall it lies in relation to a tube of peritoneum called the processus vaginalis which extends down into the scrotum. The pathway through the abdominal wall becomes known as the inguinal canal. Migration of the testis is usually completed by the time of birth. The structures linking the testis with other abdominal structures - eg: the ductus deferens and the testicular vessels and nerves - lie for part of their course within the inguinal canal. The distal part of the processus vaginalis normally becomes separated from the peritoneal cavity and forms the sac-like tunica vaginalis that surrounds most of the testis. This arrangement allows the testis to move in relation to the scrotum.
It is interesting that as the gonads descend, the kidneys ascend from their origin in the pelvic region, so that the relative positions of the gonads and kidneys are reversed.
formation of the gametes
In the developing ovary the primordial germ cells divide repeatedly and produce a population of primary oocytes surrounded by follicle cells - the primordial follicles. About 6 million of these follicles are formed by the 5th month of development, but then a process of attrition (loss) begins as some follicles begin to degenerate. By the time of birth, the number of viable follicles and oocytes has dropped to about 1/6th of the original number, and the surviving oocytes have commenced their first meiotic division. Thus in the female, gamete formation and meiosis begin before birth. The process of reduction of oocyte numbers continues after birth, and is called atresia.
The female gametes then pass into a phase of suspended development which continues at least until puberty - and for some follicles will last for many years after that. During the reproductive years which follow puberty, groups of follicles recommence their development. With each menstrual cycle, a handful of follicles begin to mature, and the oocytes within continue their first meiotic divisions. But usually only one follicle will actually ovulate in a given cycle - the others in the small group of maturing follicles become atretic.
Even at the time of ovulation, the oocyte is not yet a true ovum - it has still to complete its second meiotic division. Meiosis is only completed if fertilisation takes place - if not fertilised, the oocyte will die after about 24 hours, still halfway through its second meiotic division. If fertilisation does occur, the haploid genetic contributions of the ovum and sperm combine to form the diploid genome of the new individual, and cell is now called a zygote.
The reproductive period for women usually lasts for 30-35 years, after which the ovarian cycles cease at menopause. Even if she ovulates every month throughout her reproductive years, only 300-400 oocytes will be released, a small proportion of the 6 million or so potential ova present in the ovaries before birth.
In the male, gametogenesis does not begin until puberty, and then production of spermatozoa becomes a continuous, production-line process that continues into old age. A single ejaculate contains approximately 300 million spermatozoa.
Thus, gametogenesis follows quite different time-scales in the two sexes, and involves different numbers of gametes, even though the same fundamental meiotic events are observed in oocytes and spermatocytes.
variations in development of the reproductive system
Development towards full reproductive potential requires that decisions are made appropriately at several branch points along the developmental pathway - genetic sex, gonadal sex, internal duct system, external genitalia, brain changes, and puberty, for example. However, biologically many different combinations of features are possible. In addition, many personal and cultural pressures can affect the gender of a person.
The human embryo appears to have a predisposition to develop along the female pathway. As outlined above, the arrival of primordial germ cells carrying Y chromosomes in the gonads cause them to develop into testes, and this has knock-on effects on structures such as the two internal tubular systems linked with the gonads and the development of the external genitalia. However, this dominant influence of the Y chromosome is quite localised, and if for some reason a testis develops only on one side of the embryo, then the female-type duct system will develop on the other side. Timing in all these events is critical. To give an example, in a male embryo the genital swellings fuse in response to circulating male hormones to produce the scrotum and penis. If the male hormones are absent at this critical time, and then arrive at a later time, fusion will not occur but there will be enlargement of the clitoris and labial folds.
Here are some examples of variations in sexual development that stem from genetic and other factors:
chromosomal variants
The Y chromosome is dominant and testis-producing, but if it is accompanied by more than one other sex chromosome development of a normal testis may be prevented. Disorders of the sex chromosomes generally arise during meiosis when the gametes are being formed, but as the embryo develops there may be errors of mitosis that give rise to mosaicism - that is an embryo with two or more genetically-different cell populations. In addition to anomalies in the number of whole sex chromosomes, it is possible to have localised changes in a single sex chromosome sufficient to disrupt normal development. On the other hand, the presence of seemingly normal sex chromosomes does not guarantee development of normal gonads, and hermaphrodite people with both testicular and ovarian tissues have been identified with apparently normal chromosomal constitutions. This is a reminder of the subtle relationship between genotype and phenotype. Here are some examples of variations in sexual and reproductive development:
Turner's syndrome (44 + XO)
Each cell has only a single X chromosome. As a consequence, the person does not complete the usual sequence of puberty and retains a relatively prepubertal bodily form. Treatment may include oestrogen therapy to encourage development of secondary sex characteristics.
Triple X female (44 + XXX)
People with this condition are sometimes called 'super females' because of the extra copy of an X chromosome in each cell. However the condition may be associated with incomplete reproductive development and learning difficulties.
Klinefelter's syndrome (44 + XXY)
There is an extra X chromosome in each cell. The external genitalia are male-like, but the testes are very small and generally no spermatozoa are produced. Treatment is not usually required.
True hermaphroditism
Ovarian and testicular tissues are present in the same person. The possible combinations include an ovary with a testis, a testis with an ovotestis (a gonad with features of both ovary and testis), and ovotestis with ovotestis. The external genitalia may be male-like or female-like. The genetic make-up is usually 44 + XX, but XX/XY mosaics occur. Treatment may be given to enhance a gender preference.
Gonadal dysgenesis
The subjects are tall and female-like with normal female external genitalia. They have a uterus but no ovaries, and there is an absence of breast development. The genetic make-up is usually 44 + XY. Oestrogen therapy may be given to enhance female secondary sexual characteristics.
Masculinisation in the female
Those affected are genetically female. The degree and type of masculinisation depends on whether the causative factors exerted their effect in prenatal life, childhood, or in adult life. Masculinisation that begins before birth modifies development of the genital organs, and the degree of masculinisation may be sufficient to give rise to mistaken identification of sex at birth. Prenatal masculinisation may be due to an adrenal or ovarian dysfunction in the mother or the fetus. For example, congenital adrenal hyperplasia is due to inadequate synthesis of the hormone cortisol by the adrenal glands of the fetus. As a result there is hypertrophy of the clitoris and persistence of the urogenital sinus. If untreated after birth, the masculinisation continues and there is precocious growth of pubic hair, deepening of the voice, no breast development, diminished growth, and infertility. Treatment is by administering cortisone. Masculinisation can also be caused before birth by male-type hormones reaching the baby from the mother. For example, some synthetic progestins that were given to mothers to maintain pregnancies that threatened to miscarry were found to produce masculinisation in female offspring. Alternatively, androgens may be produced within the mother because of a pathological condition and passed across to the fetus where they cause masculinisation. Clitoral enlargement and fusion of the labia majora may occur.
Feminisation in males
Feminisation of a genetic male may occur during fetal life. The main type is testicular feminisation. It is due either to very early failure of the fetal testes to develop, or a defective response of tissues to fetal androgens. Patients with the testicular feminisation syndrome have female-type external genitalia and body form. At puberty they develop breasts but do not grow pubic or axillary hair and do not menstruate. The vagina is absent or poorly developed, there is no uterus, and the testes are undescended. Often, no treatment is required since the person is physically and psychologically female, even though the karyotype is 44 + XY.
There is another condition which may lead to mistaken identification of sex at birth: hypospadias. This is due to failure of output of androgens by the fetal testes at the time of closure of the urethral folds, with the result that the urethra opens beneath the penile shaft rather than at the tip of the glans penis.